
Frank van Hertrooij
Scivolemo 30 Aprilo 2021
Pliaj artikoloj
Kiam mi estis infano, la BBC elsendis Brita-Kenja infana televidserio pri kial la bestoj estas kiaj ili estas. Mi tro maljunis por ĝi, sed mi vidis kelkajn epizodojn kiam mi malsanete restis hejme de la lernejo kaj la televido montris nenion alian. Mi ne certas pri kial mi tiel klare memoras la serion, sed mi kulpigas la artstilon. Ĝi estis bunta kaj elstara, kaj la bestoj havis belajn desinojn*. La stilo estis bazita sur la Tingatinga arto el la oriento de Afriko, de kie devenis ankaŭ la fabloj kiuj inspiris la epizodojn. Ĝi estis taŭga, ĉar afrikaj bestoj estas oftaj temoj en Tingatinga arto, kaj per siaj akraj kontrastoj la stilo bone montras la varion de iliaj ŝablonoj. Nun mi estas plenkreskulo kaj mi daŭre scivolas pri bestoj. Kvankam la fabloj nun lasis spacon por scienco, tio ne igas la bestojn malpli belaj. Fakte, lerni iom pli pri la origino kaj meĥanismoj malantaŭ bestaj desinoj igis min aprezi ilin eĉ pli. Verdire, kio estas pli bela ol krei tutan beston el unu ĉelo?
*Kiam mi uzas la vorton “desino” en ĉi tiu afiŝo, mi parolas pri (preskaŭ)ripetataj formoj en spaco aŭ tempo. Por ĉi tiu koncepto ekzistas pluraj vortoj, kiuj ĉiuj havas avantaĝojn kaj malavantaĝojn. Atendante oficialan respondon de la AdE, mi uzas la neologismon “desino” laŭ la persona rekomendo de unu membro de la AdE.

Malsamaj ĉeloj, sama DNA
Unu el la nombroj kiujn scienckomunikistoj ŝatas krii estas ke 99% de la DNA de homoj kaj ĉimpanzoj estas identa. Kvankam la nombro povas esti iomete malpli alta depende de kiajn mutaciojn oni enkalkulas, restas fakto ke homoj kaj ĉimpanzoj ne tiom malsamas. Eble tio ne estas surprizo kiam temas pri niaj plej proksimaj parencoj. Sed ankaŭ kiam oni prenas pli malproksiman beston kiel muson, oni daŭre trovos ke pli ol duono de la DNA estas identa; en kelkaj gravaj genoj eĉ 99%. Kiam oni legas ĉi tiajn nombrojn sen multe da kunteksto, ili ne signifas multe. Klare ne ĉiuj genoj ŝanĝiĝas same rapide, sed kial? Kiom da mutacioj necesas por krei videblan ŝanĝon?
Kelkaj genoj estas tiel antikvaj ke ili apenaŭ malsamas inter homoj kaj bakterioj. Temas ĉefe pri genoj kiuj havas rolon en la reproduktado de ĉeloj. Se okazus mutacio en tia geno, la ĉelo verŝajne ne plu povas reprodukti sin, do la mutacio ne transdoniĝas. Tiaj genoj estas ”konservataj”. Kiam oni rigardas nur bestajn ĉelojn, oni vidas multajn el tiaj evolue konservataj genoj, ĉar bestaj ĉeloj funkcias esence same. La grandaj diferencoj inter mamuloj ofte ne esprimiĝas je la ĉela nivelo, sed la hista aŭ organa nivelo. Oni povas demandi sin kiel tio eĉ eblas. Kiel DNA-mutacio povas fari ŝanĝon en nur unu histo se ĉiu ĉelo en la korpo havas la saman DNAn? Eble la genaro ne estas recepto kiun la ĉelo legas de la komenco ĝis la fino, sed manlibro el kiu ĝi povas preni nur la koncernajn informojn.
Malsataj bakterioj
Eĉ simpla organismo kiel bakterio havas multajn genojn. Ĝi povus uzi ĉiujn el tiuj genoj samtempe, sed tio estus malefika en la plej bona kazo kaj letala en la plej verŝajna kazo. Fabriki proteinojn kostas multe da energio kaj materialoj, do se bakterio povas krei nur la proteinojn kiujn ĝi bezonas en la momento, ĝi havas avantaĝon. Tial aperis multaj reguligaj meĥanismoj jam frue en evoluo. Tiuj meĥanismoj similas iomete al la logiko de komputilaj programoj en la senco ke partoj de la kodo nur estas legataj kiam specifa kondiĉo veras. La diferenco estas ke la sistemo estas tute molekulaj.
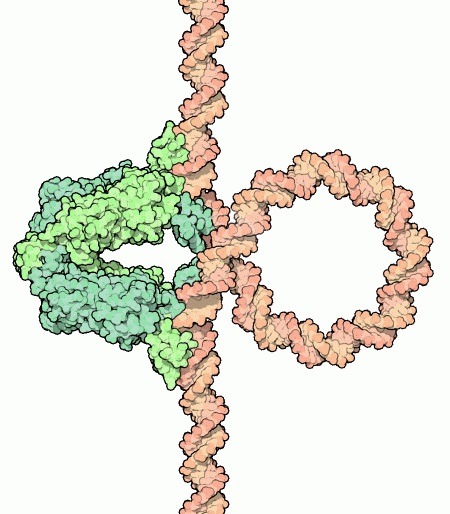
Unu el la plej famaj ekzemploj de genregulado estas la Lac-operono de la bakterio E. coli. Tio estas grupo de tri genoj, LacZ, LacY kaj LacA, kiuj necesas por la sorbado kaj digestado de laktozo. Kutime la bakterio ne bezonas tiujn genojn, ĉar ĝia preferata nutraĵo estas glukozo, sed nur laktozo estas alirebla, ĝi ŝanĝu sian strategion. Du gravaj molekuloj en tiu reguliga strategio estas RNA-polimerazo kaj la Lac-supremanto. RNA-polimerazo estas molekula kopiilo, kiu transskribas genojn de DNA al RNA, por ke ili povu esti tradukataj al proteinoj. La Lac-subpremanto estas proteino kiu ligas sin al la DNA tuj antaŭ la genoj, kaj blokas la vojon por RNA-polimerazo. Tio signifas ke la genoj ne povas esti uzataj kiam la Lac-supremanto estas ligita. La Lac-supremanto en si mem estas interesa kaj kompleksa molekulo, kiu konsistas el tri gravaj partoj. Unu flanko estas strukturita tiel ke ĝi povas interagi kun la formo kaj ŝarĝdistribuo de specifa sekvenco en la DNA. Tiel ĝi scias ekzakte kie ĝi ligu sin. La alia flanko de la proteino povas ligi sin al tri kopioj de la sama proteino, por ke ili povu kunigi siajn fortojn. Plej grava estas la kerno tamen. Tio estas la plej granda parto de la proteino, kiu povas rekoni alolaktozon, molekulo kiu kreiĝas dum la digestado de laktozo. Alolaktozo ligas sin al la kerno de la Lac-supremanto kaj ŝanĝas ties formon tiel, ke ĝi ne plu povas ligi sin al la DNA. Tiel la digestado povas pludaŭri ĝis la alolaktozo estas tute digestita kaj la signalo malaperas. Tiam la digestaj proteinoj ne plu necesas kaj Lac-supremanto denove ligas sin al la DNA por bloki la transskribadon. Estas perfekta ekzemplo de kiel proteinoj povas reguligi kiel genoj esprimiĝas por adapti sin al ŝanĝoj en la medio. Nun la granda demando estas ĉu la samo veras pri la homa ĉelo.
La koordinata sistemo de muŝoj
Homaj genoj estas similaj al bakteriaj genoj en la senco ke ili havas genetikajn ŝaltilojn. Ekzistas mallongaj sekvencoj antaŭ la geno al kiuj proteinoj povas ligi sin por bloki aŭ plifortigi la esprimadon. Tio estas tre interesa sistemo, ĉar la ŝaltiloj de tute malsamaj genoj povas esti identaj. Tiel unu proteino povas influi plurajn genojn. Aldone, tiuj proteinoj mem venas de genoj, kiuj estas same strikte regulataj. Ekestas ŝtuparo de proteinoj, kiuj reguligas la genojn de aliaj reguligaj proteinoj, iĝante pli kaj pli specifaj. La plej specifaj reguligaj proteinoj laboras je la nivelo de individuaj ĉeloj, kreante tre fajnajn detalojn. Dume, la tre ĝeneralaj laboras je la nivelo de tutaj organoj kaj decidas aferojn kiel ”La okuloj estu tie!” aŭ ”Tiuj vertebroj havu ripojn kaj tiuj ne!”
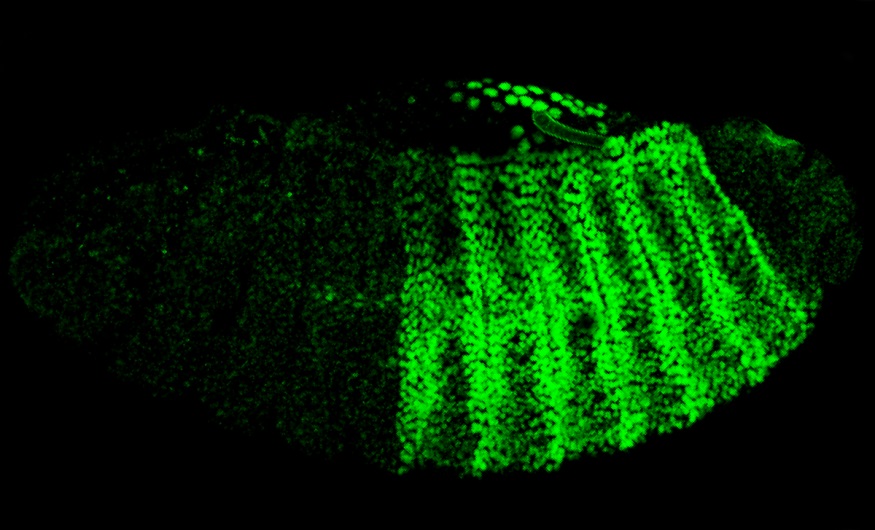
Oni povas sekvi kiel reguligaj proteinoj kreas malsamajn strukturojn en la embrio. La plej simpla maniero por fari tion estas per GFP, verda fluoreska proteino, kiu sorbas bluan lumon kaj ellasas verdan lumon. En laboratoriaj eksperimentoj oni povas fiksi GFP-proteinojn al antikorpoj kiuj rekonas tiujn reguligajn proteinojn, kaj vidi en kiuj ĉeloj la antikorpoj abundiĝas per mikroskopo. Farante tion por pluraj reguligaj proteinoj kaj dum longa tempo, oni povas vidi kiel la proteinoj aperas kaj malaperas dum la malsamaj fazoj de la embria disvolviĝo kaj kiel ekestas kompleksaj desinoj. La GFP koncentriĝas en strioj kaj makuloj kun akraj limoj, kaj se oni bone sekvas tiujn desinojn dum la disvolviĝo, oni vidas ke ili korespondas al la lokoj de estontaj organoj. Unu el la plej interesaj demandoj kiujn oni povas demandi estas kiel ŝajne simetria ovoĉelo povas iĝi tiel multe malpli simetria.
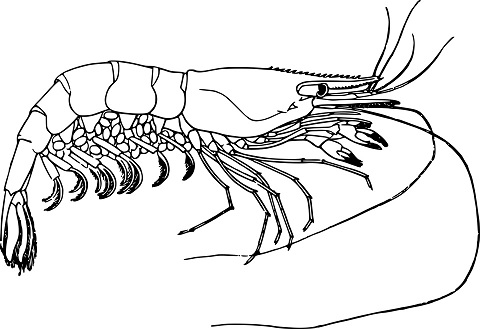
Por esplori tiun demandon, mi ŝatus paroli pri la nigraventra drozofilo, la bananmuŝo, ĉar ĝi estas unu el la plej multe uzataj laboratoriaj bestoj. Ĝia embria disvolviĝo ne estas tute sama kiel tiu de homoj, sed kio gravas estas la ĝenerala principo. Kiel ĉelo scias kie ĝi estas en la embrio? La sekreto estas ke la ovo ne estas tute simetria. Longe antaŭ la fertiliĝo, kiam la patrino kreas la ovon en sia korpo, ŝi enmetas genojn en la formo de RNA-molekuloj, kiuj estas fiksitaj al unu flanko de la ĉelo. La unua el tiuj genoj kiujn oni malkovris nomiĝas ”bicoid”. Ĝi ekzistas nur en muŝoj, sed aliaj insektoj faras la saman per aliaj genoj. Nur post la fertiliĝo, bicoid liberiĝas kaj povas esti tradukata al proteino, kiu difuziĝas tra la embrio. La bicoid-proteino estas morfogeno. Ĝi estas molekulo kies distribuo difinas la strukturon de la embrio. La koncentriteco de bicoid malkreskas laŭ la longo de la embrio kaj tiel kreiĝas unu el la unuaj desinoj de la embrio. Ĝi eble ne sonas kiel tre interesa desino, sed ĝi estas gravega. Surbaze de la koncentriteco de bicoid, ĉelo povas scii kiom longe for de la antaŭo de la estonta kapo ĝi estas. Kune kun aliaj proteinoj kiel nanos, kiu esprimiĝas en la malantaŭa flanko de la embrio, ĝi kreas la unuan akson de molekula koordinata sistemo. Aliaj genoj uzas tiun koordinatan sistemon por krei pli detalajn desinojn. La gap-genoj, ekzemple, difinas la limojn inter la malsamaj segmentoj de la embrio. Ili esprimiĝas nur tie kie la koncentritecoj de morfogenoj estas ekzakte ĝustaj, kreante zonojn.

Membroj
Malpli abstraktaj ol genoj kaj proteinoj estas bestoj. Kiel ni povas vidi la efikojn de reguligaj genoj en ili? Multaj partoj de niaj korpoj fakte estas malsame regulitaj versioj de la sama organo. Ekzemplo de tio estas niaj membroj. Niaj kruroj kaj brakoj aspektas sufiĉe malsimilaj, sed tio ne veras por niaj plej proksimaj parencoj, la nehomaj simioj. Se oni rigardas kiel orangutano moviĝas inter arboj, oni rimarkas ke ili uzas ankaŭ la piedojn por preni branĉojn. Iliaj piedoj estas aldona paro de manoj kun funkciantaj dikfingroj. Homoj ne ofte grimpas en arbojn kaj certe ne uzas la piedojn tiel. Niaj manoj povas preni objektojn kaj eĉ skribi kaj tajpi, dum niaj piedoj estas bonegaj por longdistanca kurado. Anstataŭ havi du funkciojn, manipuladon kaj transporton, niaj membroj specialiĝis.

Eĉ pli da diverseco troveblas inter artropodoj, kiuj havas multe pli da membroj. Kelkaj havas eĉ pli da membroj ol TEJO. Kiam oni rigardas salikokon, oni vidas kelkajn specojn. Ili troviĝas en grupetoj laŭ la longo de la besto. Kelkaj el ili estas por marŝado, aliaj por naĝado, kaj tiel plu. Tre verŝajne la membroj de iliaj prauloj estis multe pli similaj, sed kelkaj specialiĝis. Se oni studas artropodojn genetike, oni vidas ion eĉ pli belan kaj universalan. Ne nur la kruroj, sed ankaŭ la antenoj, buŝo-partoj kaj multe pli; ili ĉiuj estas specialigitaj membroj. Tio reflektiĝas en la genetiko. La korpo de artropodo konsistas el multaj segmentoj, kiuj ĉiuj povas havi unu paron de subaj membroj. Tio facile videblas en centpieduloj, sed malpli en ekzemple insektoj kaj araneoj. Kvankam ankaŭ tiuj grupoj havas multajn segmentojn, ili kunfandiĝas kaj ŝanĝiĝas dum la larvo maturiĝas. La identeco de la segmentoj kaj iliaj membroj estas multe influataj de la tiel nomataj hox-genoj. Ili estas familio de genoj kiuj esprimiĝas malsame laŭ la longo de la embrio. Ĉi tiu sistemo estas tre facile tajlorebla, ĉar la segmentoj iusence povas evolui sendepende. Ili kaŭzas ke mutacio influas nur kelkajn segmentojn anstataŭ la tutan korpon, kaj ke la identeco de segmento povas ŝanĝiĝi per regulado de genoj anstataŭ la grada, malrapida evoluo de la genoj mem. Eble la fakto kiu plej bone montras la utilon de hox-genoj estas la fakto ke ili estas antikvaj. Ili disvolviĝis longe antaŭ la Kambrio kaj hodiaŭ troviĝas en esence ĉiuj bestoj krom sponguloj, do ili certe funkcias bone.
Kompare al ekzemple krustaceoj, kies membroj estas kiel svisa tranĉilo, insektaj membroj estas relative tedaj. Ili multe varias de specio al specio, sed kun nur tri paroj oni ne povas facile distribui la taskojn. La genetika kialo estas simpla. La hox-geno antennapedia aktivigas la genojn kiuj necesas por la kreado de kruroj. Abdonimal-A subpremas la formiĝon de kruroj. Tio signifas ke kuroj nur formiĝas en la tri segmentoj kiuj esprimas antennapedia, sed ne abdominal-A. Havi malmulte da membroj kreas limigojn, sed ankaŭ oportunojn. Pli da membroj estas pezaj, kostas pli da energio kaj ili povas esti en la vojo. Tio estas problemo sur la grundo, sed eĉ pli en la aero.
Flugiloj
Pli imponaj ol la kruroj estas la flugiloj. Insektoj estis la unua grupo de bestoj kiuj ekflugis (100-200 milionoj da jaroj antaŭ vertebruloj) kaj tio verŝajne estis grava faktoro de ilia sukceso. Hodiaŭ pli ol miliono da specioj estas konataj kaj granda plejparto flugas. Interesa observo kiun oni povas fari estas ke muŝoj kaj kuloj (dipteroj) havas nur du flugilojn kaj tio estas surprize rara kondiĉo. Libeloj klare havas kvar grandajn samaspektajn flugilojn, sed ankaŭ vespoj kaj abeloj, termitoj, kaj formikoj. Interesa komparo al muŝoj estas papilioj. Ili havas du parojn de flugiloj, sed la malantaŭa paro estas malpli granda kaj povas aspekti sufiĉe malsimile. Se oni rigardas muŝon, serĉante specialigitajn flugilojn, oni trovas ion malsaman: halterojn.

Halteroj estas malgrandaj organetoj kiuj funkcias iom kiel giroskopo kaj provizas informojn pri la flugado. Ili troviĝas tie, kie la malantaŭa paro de flugiloj devus troviĝi. Genetike tio estas klarigebla per la hox-geno ultrabithorax. Ĝi funkcias per bloki aliajn genojn kiuj necesas por la disvolviĝo de flugilo. Oni povas konfirmi tion per genetike modifi muŝon tiel, ke ĝi ne plu esprimas ultrabithorax. La rezulto estas muŝo kun du paroj de funkciantaj flugiloj. Ankaŭ en papilioj la diferencoj inter la du paroj de flugiloj estas kaŭzataj de ultrabithorax, sed ĝi influas aliajn genojn ol en muŝoj. Se oni malaktivigas ultrabithorax en papilioj, la du flugiloj havos multe pli similan formon kaj desinon.
La malo okazas en unu el la plej grandaj grupoj de insektoj: koleopteroj. En tiu grupo, la malantaŭa paro de flugiloj estas malplej specialigita. Anstataŭe, la antaŭa paro iĝis elitro. Tio estas malmola kovrilo, kiu protektas la verajn flugilojn. Kvankam la rezulto estas mala, ankaŭ ĉi tiu kondiĉo povas esti klarigata de ultrabithorax. Ĉi-kaze la “programo” por krei flugilojn ŝanĝis al programo por elitro. Per bloki genojn en tiu elitra programo, oni povas krei flugilon simila al tiuj de aliaj insektoj. Tiu blokado estas farata de ultrabithorax kaj okazas nur en la malantaŭa flugilo.

Desinoj de Turing
Ĉiuj ekzemploj de bestaj desinoj kiujn mi montris ĝis nun temis pri koordinata sistemo. Ĉeloj scias kion ili faru, ĉar ili mezuras la koncentritecon de malsamaj morfogenojn. Ĉu tio sufiĉas por klarigi ĉiujn striojn, makulojn kaj aliajn artaĵojn kiuj videblas en la naturo? Verŝajne ne. Ekzistas ankaŭ aliaj meĥanismoj kiuj verŝajne havas gravan rolon en la desinoj de bestoj, kaj unu el la plej gravaj estas desinoj de Turing. La elpensinto, Alan Turing, estas pli konata pro sia kerna rolo en la malĉifrado de la naziaj ”Enigma”-ĉifriloj, la teoria fundamento de modernaj komputiloj, kaj la kruelaĵojn kiujn li spertis pro sia samseksemo. Por ĉi tiu rakonto, tamen, gravas publikaĵo lia el 1952. Lia matematika fono reflektiĝas en la abstrakteco de la publikaĵo, kaj la diferencialaj ekvacioj povas fortimigi nematematikistojn. Eble estas tial ke la publikaĵo nur kovriĝis de polvo dum jardekoj ĝis ĝi finfine estis remalkovrita.

La publikaĵo, The Chemical Basis of Morphogenesis (La ĥemia bazo de morfogenezo), temas pri kiel simpla sistemo konsistanta el du difuziĝantaj molekuloj (aktiviganto kaj inhibanto) povas krei la plej kompleksajn desinojn. La sekreto estas ke la molekuloj influu unu la alian. La aktiviganto stimulu la fabrikadon de si mem kaj de la inhibanto, dum la inhibanto bloku la fabrikadon de la aktiviganto. Tia sistemo ne estas tute fremda al biologio. En la suba grafikaĵo, ekzemple videblas modelo de la populacioj de predoj kaj predantoj. Teorie tiuj populacioj influas unu la alian same kiel la aktiviganto kaj inhibanto. Predoj pligrandigas sian propran populacion per seksumado kaj tiun de la predanto per esti bongusta manĝaĵo. La predantoj dume, blokas la kreskadon de la preda populacio per ĉasado. Se la predoj multobliĝas, ankaŭ la predantoj multobliĝas, ĝis la kvanto de predoj komencas malkreski kaj ĉio inversiĝas. La rezulto estas onda desino tra la tempo. Io simila povas okazi en Turing desino. La koncentritecoj de la molekuloj estas kiel la populacioj de la predoj kaj predantoj, sed anstataŭ tra tempo, la onda desino aperas laŭ la longo de la embrio, ĉar la molekuloj difuziĝas de unu parto de la embrio al alia. Poste, la genoj kiuj necesas por krei kolorojn povas aktiviĝi nur tie, kie la koncentritecoj de la aktiviganto kaj inhibantoj estas ĝustaj.
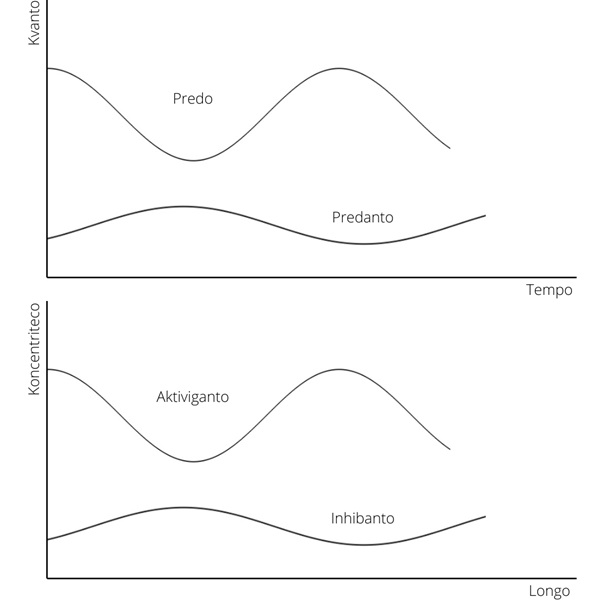
Se la difuziĝado okazas laŭ la longo de iu korpoparto, formiĝas strioj. Se ĝi okazas dise el centra punkto, makulo formiĝas. La rapideco per kiu la molekuloj difundiĝas povas influi kiom grandaj tiuj makuloj estas kaj ĉu ili povas konektiĝi. Tia sistemo ne nur utilas por pentri la haŭton de besto, sed ankaŭ por aliaj desinoj kiel la loko de haroj aŭ plumoj sur la haŭto, aŭ eĉ la pozicio de fingroj en mano. Pruvi ke iu desino estas la rezulto de desino de Turing ne estas facila tasko, sed la desinoj kreitaj de komputiloj estas tiel similaj al la naturo, kaj oni eĉ trovis ekzemplojn de molekuloj, kiuj interagas en la ĝusta maniero.
Estas klare ke la kreado de besto ne estas simpla procezo. Por eĉ la plej simplaj bestoj necesas multaj genoj, kompleksaj hierarĥioj, gradientoj, aktivigantoj, inhibantoj kaj pli, sed la listo ne estas senfina. Same kiel artisto povas per limigitaj materialoj, iloj kaj informoj krei senliman varion, la naturo kreis ĉiujn bestojn kiujn ni konas el modesta praulo. Ni povas aprezi la artaĵojn de la naturo pli bone se ni komprenas ilin. Sed, kiel artisto, la naturo ne malkovros siajn sekretojn sen laborigi nin.
Plua legado:
Carroll, S. B. (2006). Endless Forms Most Beautiful (1st ed.). Ww Norton & Co.
PDB-101: Molecule of the Month: lac Repressor. (n.d.). Retrieved April 30, 2021, from http://pdb101.rcsb.org/motm/39
Tomoyasu, Y., Wheeler, S. R., & Denell, R. E. (2005). Ultrabithorax is required for membranous wing identity in the beetle Tribolium castaneum. Nature, 433(7026), 643–647. https://doi.org/10.1038/nature03272
Licencoj por la bildoj:
Tingatinga – flowcomm/Tingtinga Arts Cooperative Society: https://www.flickr.com/photos/flowcomm/49186344216
Lac-supremanto – David S. Goodsell/RCSB PDB: http://pdb101.rcsb.org/more/how-to-cite
Drozofilo kun GFP – Caitlin Sedwick: https://commons.wikimedia.org/wiki/File:Stage-14_Drosophila_embryo_expressing_AbdA_and_Exd,both_fused_to_one_half_of_the_fluorescent_protein_Venus-_journal.pbio.1001349.g001.png
Orangutano – Thomas Quine: https://commons.wikimedia.org/wiki/File:How_to_peel_a_banana_with_your_foot_(26443349170).jpg
Tipolo – ©Entomart: https://commons.wikimedia.org/wiki/File:Tipula_oleracea01.jpg
Desino de Turing – Jonathan McCabe: https://www.flickr.com/photos/jonathanmccabe/4309502565
{kind=link}
.jpg){kind=link}
{kind=link}